
Oliver Gluchs Welt der Fleischfressenden Pflanzen oder: "Was Sie schon immer einmal über Fettkraut wissen wollten" |
| Impressum |

| Startseite | Vorkommen | Beutetiere | Arten | Pflanzenkauf | Andere Karnivoren | Infos über Karnivoren | über mich |
|
Pinguicula casperiana Crespo, Martínez-Azorín, Alonso & Sáez (2020)Aus
der
Serranía de Cuenca, einem Kalkstein-Gebirge in der
zentralspanischen Provinz Castilla-La
Mancha östlich von Madrid, wurden in der
Vergangenheit schon an mehreren Stellen
Fettkrautpopulationen gefunden. Das Gebirge durchziehen
große, zusammenhängende Waldgebiete, vorwiegend aus den
Baumarten Pinus nigra (Schwarzkiefer), Quercus
faginea (Portugiesische Eiche) und Quercus
ilex (Steineiche) bestehend. Die Landschaft ist
auch geprägt von tiefen Tälern und Schluchten mit
speziellem Mikroklima, eine der bekanntesten ist die Hoz
de Beteta.
Bisher wurden diese Fettkrautpopulationen aus dieser Gegend immer P. grandiflora oder P. dertosensis zugeordnet. 2020 kamen Wissenschaftler der Universität Valencia rund um den spanischen Botaniker Manuel Crepso bei Untersuchungen von verschiedenen spanischen Fettkrautpopulationen zu dem Schluß, dass es sich bei den Sippen aus der Serranía de Cuenca um eine eigenständige Art handelt und beschrieben die Pflanzen unter dem Namen Pinguicula casperiana (zu Ehren des deutschen Botanikers Jost Siegfried Casper, der mit seiner Monographie zur Gattung Pinguicula in den 1960er Jahren ein Standardwerk über die damals weltweit bekannten Fettkrautarten verfasste). Allerdings zeigen die molekulargenetischen Untersuchungen eine starke Verwandschaft zu P. dertosensis und die beschriebenen morphologischen Unterschiede zu P. dertosensis sind nur minimal. Die Behauptung von Crespo et al. P. dertosensis würde nur eine karnivore Blattform aufweisen, während P. casperiana anisophylle Sommerblätter (zwei unterschiedlich geformte karnivore Blatttypen) bildet, spiegelt nicht ganz die Realität. Es gibt Populationen von P. dertosensis, die deutliche Unterschiede von karnivoren Blatttypen aufweisen, während bei anderen Populationen ein Unterschied nur in der Blattlänge sichtbar wird. Daher kann die Eigenständigkeit von P. casperiana durchaus kritisch betrachtet werden. Die Pflanzen wachsen an dauerhaft feuchten oder wasserüberrieselten vertikalen Kalksteinwänden, die meist schattig gelegen sind. Bisher wurden Populationen dieser Art zwischen 950 und 1550 m ü. M. gefunden. 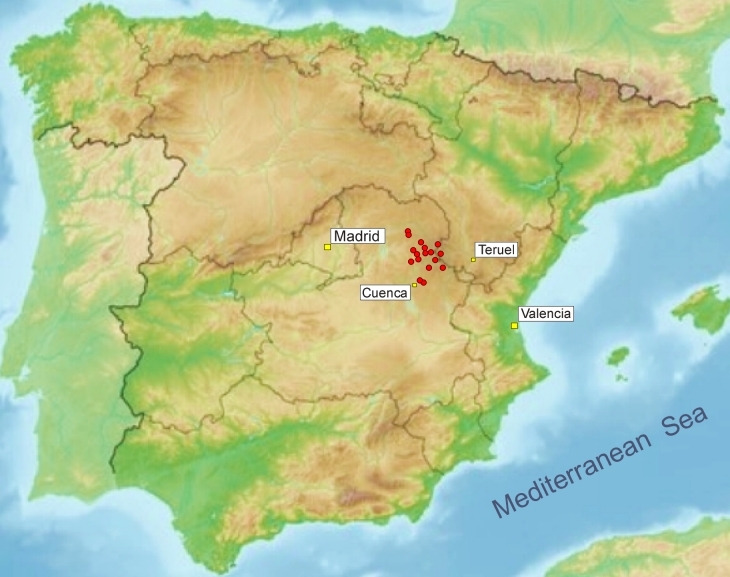 P.
casperiana
überdauert
die
kalten Wintermonate in Form eines Winterknospe, die auch
als Hibernaculum bezeichnet wird. Diese Winterknospe
wird schon im Sommer ausgebildet, vielleicht als
Reaktion auf die recht trockenen Bedingungen Ende des
Sommers und als Schutz vor den sehr kalten Bedingungen
während des Winters in den Höhenlage der Serranía de
Cuenca. Im März/April treiben die ersten karnivoren
Blätter aus. Laut den Autoren ist die Art
subheterophyll, was bedeutet, dass die Art 2
verschiedene karnivore Blatttypen ausbildet, die sich
laut den Autoren aber nur in ihrer Größe unterscheiden.
Die Blätter der "Frühjahrsrosette", die sich vom
Austrieb ium Frühjahr bis etwa zur Blütenknospenbildung
erstreckt, werden 2 bis 5 cm lang und bis zu 2 cm breit,
sind länglich-verkehrt-eiförmig und laufen am Blattende
leicht spitz bis spitz zu. Etwa ab dem Zeitpunkt
der Blüte werden die "Sommerblätter", die mit 3,5 bis 7
cm länger und mit 1 bis 2,8 cm auch etwas breiter sind
und eine länglich-verkehrt eiförmig bis
länglich-elliptische Form haben. Die Blattenden sind
leicht spitz zulaufend bis rundlich. Beide Blattformen
sind auf der Oberseite dicht mit Drüsenhaaren besetzt
und der Blattrand ist nach oben gebogen.
      Die
Hauptblühperiode
von P. casperiana
erstreckt sich von Mitte April bis Mitte Mai. Dabei
können 1 bis 8 Blüten pro Pflanze ausgebildet werden,
wobei der Blütenstiel eine Länge von 4 bis 8 cm,
selten bis 10 cm erreicht. Die Blütenstielbasis ist
hellgrün, danach wird der Blütenstiel rotbraun, um dann
zur Blüte hin eine dunkelviolette Farbe anzunehmen. Der
Blütenkelch ist zweilippig, die Kelchblätter sind meist
dunkelviolett oder braunviolett gefärbt und auf der
Oberseite der Kelchblätter sind vereinzelt gestielte
Drüsenhaare vorhanden. Die Kelchblätter haben eine
länglich-lanzettliche Form mit meist abgerundetem oder
abgeflachtem Ende. Die Blütenkrone ist ebenfalls
zweilippig, wobei Ober- und Unterlippe einen Winkel
zwischen 70 und 120 Grad bilden. Die Petalen der
Oberlippe sind 6 bis 7 mm lang und 5 bis 6 mm
breit, verkehrt-eiförmig, überlappen sich nicht und
sind leicht nach hinten gebogen. Die Farbe der oberen
Kronblätter ist lila bis violett. Die Kronblätter der
Unterlippe sind größer, die seitlichen Kronblätter sind
8 bis 10 mm lang und 4 bis 6 mm breit, der Mittellappen
mit 9 bis12 mm länger und mit 8 bis 10 mm auch breiter.
Die Form der Kronblätter ist rundlich bis länglich
verkehrt-eiförmig. Die Farbe der Unterlippenkronblätter
ist außen lila bis violett, die Basis ist je nach Blüte
unterschiedlich groß weiß gefärbt. Die Oberseite
der Kronblätte ist dicht mit weißlich-violetten Härchen
besetzt. Die trichterförmige Kronröhre ist violett
gefärbt mit parallel verlaufenden violetten Adern und
der Kronröhreneingang ist relativ groß und breit. An die
Kronröhre schließt sich der braun-violett gefärbte Sporn
an, der bis zu 13 mm lang wird (selten bis zu 16 mm) und
gerade bis leicht gebogen ist.
Laut den molekulargenetischen Untersuchungen von Crespo und Kollegen scheint die Art am engsten mit P. dertosensis verwandt zu, allerdings sind die morphologischen Unterschiede nicht sehr groß. P. casperiana besitzt 2n = 64 Chromosomen.    Da
es
sich bei P. casperiana um eine
temperierte Art handelt, die mit einer Winterknospe
überwintert, muss man ab dem Spätherbst dafür sorgen,
dass die Temperaturen nachts bei unter 10 °C liegen. Das
Überwintern während der Wintermonate sollte bei
Nachtemperaturen von nicht höher als 3 und 5° C
erfolgen. Bei höheren Temperaturen ebsteht die Gefahr,
dass die Pflanzen wieder austreiben und dann meist
absterben. EIne Kultur im Freiland unter
mitteleuropäischen Bedingungen ist auch möglich, da die
Art frostresistent ist. Für die Kultur ist ein
luftdurchläßiges Substrat zu bevorzugen.
Insbesondere während der Ruhephase ist Staunässe zu
vermeiden, da sonst die Gefahr besteht, dass die
Winterknospen von Pilzen befallen werden und
verfaulen.
|